Tüm omurgalılarda kardiyak apeks, mide ve dalak sola, karaciğer ve safra kesesi sağa yerleşerek karakteristik asimetri gösterir. Sol-sağ aks anormallikleri ve lateralite defektleri insanlarda 1/7000-10000 sıklığında görülür. Embriyo saat yönünün tersine bükülürken diğer organların ve tüm vücudun sol-sağ asimetrisi meydana gelir. Sol-sağ asimetrisi embriyonun gelişiminde erken dönemde organların oluşumundan önce olmaktadır. Sol-sağ asimetrisini düzenleyen yollar kardiyak gelişimde de etkilidir.
Tüm omurgalılarda kardiyak apeks, mide ve dalak sola, karaciğer ve safra kesesi sağa yerleşerek karakteristik asimetri gösterir. Sol-sağ aks anormallikleri ve lateralite defektleri insanlarda 1/7000-10000 sıklığında görülür. Embriyo saat yönünün tersine bükülürken diğer organların ve tüm vücudun sol-sağ asimetrisi meydana gelir. Sol-sağ asimetrisi embriyonun gelişiminde erken dönemde organların oluşumundan önce olmaktadır. Sol-sağ asimetrisini düzenleyen yollar kardiyak gelişimde de etkilidir.
Sol-sağ asimetrisini düzenleyen yollardaki karışıklıklar heterotaksi olarak bilinen lateralite defektine, anatomik uyuşmazlığa, yapıların kaybına (aspleni), simetri ya da lateralizasyon bozukluğuna, simetrik embriyonik organların gerilememesine (persistan sol superior vena kava) ve yetmezliğine yol açar.
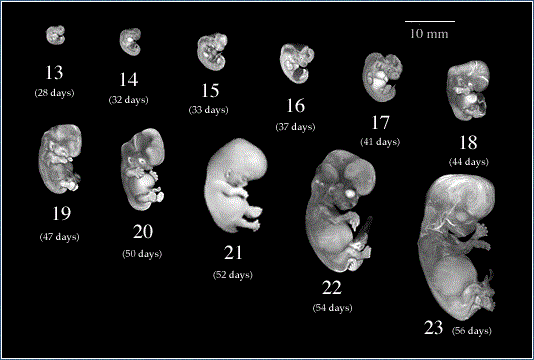
İnsan gelişimi fertilizasyon (döllenme) ile başlar ve doğuma kadar çeşitli dönemlerden geçer. Bunlar:
- Fertilizasyon (döllenme), zigotun yarıklanması, blastosist oluşumu (1. HAFTA)
- Bilaminar embriyonik disk ve koryonik kese oluşumu (2. HAFTA)
- Germ tabakalarının oluşumu, doku ve organların farklılaşmaya başlaması (3.HAFTA)
- Organogenezis dönemi (4-8. HAFTALAR)
- Fetal dönem (9. HAFTADAN DOĞUMA KADAR)
Üç germ tabakasının her biri (ektoderm, mezoderm ve endoderm) özel doku ve organları oluşturur. Epiblastların çizgisel bant halinde kalınlaşmasıyla oluşan opasite, primitif çizgi olarak bilinir ve embriyonik diskin dorsalinde, orta çizgi üzerinde ve kaudalde yerleşiktir. Primitif çizgi, kaudal ucuna hücrelerin eklenmesiyle bu yönde uzanırken, kranial ucunda hücre çoğalmasıyla primitif düğüm (Hansen’s nod) oluşur.
Bu sırada primitif çizgide primitif oluk gelişir. Primitif oluk, primitif düğümde
oluşan bir çöküntü ile devam eder ve bu çöküntüye primitif çukur denir. Primitif çizgi
belirir belirmez embriyonun kaudal ekseni, kranial ve kaudal yönleri, dorsal ventral yüzü,
sağ ve sol tarafı belirlenir.
Primitif çizginin derinindeki hücreler ayrılarak mezenşim ya da mezoblast denilen gevşek bağ dokusunu oluştururlar. Mezenşimal hücrelerin bir kısmı ise hipoblast tabakasını işgal eder. Bu tabakaya endoderm denir. Bu iki hücre tabakasının en üstteki tabakasına ektoderm adı verilir.
Paraksiyel mezoderm farklılaşarak somitleri bunlar da aksiyel iskeletin (kafa kemikleri, omurga, kostalar ve sternum) büyük kısmını oluştururlar. Lateral mezoderm içinde intraembriyonik sölom (vücut boşluğu) oluşur. Bu boşluk daha sonra peritoneal boşluk gibi diğer vücut boşluklarının kökenini oluşturur. Gastrulasyon dönemi, kan damarları, kanın oluşması ve koryon villuslarının oluşumunun tamamlanması ile sonlanır.
Kalbin primordiumu ilk olarak 18. günde belirir ve 22-23. günde atmaya başlar. Kardiyojenik alanda perikardiyal sölom’un ventralindeki splanknik mezenşimal hücreler toplanırlar ve iki longitudinal hücresel kardiyak primordiayı–anjioblastik kordonları yaparlar. Bu kordonlar kanalize olarak iki adet ince duvarlı endokardiyal kalp tüplerini oluştururlar. Lateral embriyonik katlanmalar oluştuğunda endokardiyal tüpler birbirine yaklaşır ve tek bir endokardiyal tüp yapmak üzere birleşir. Endokardiyal tüplerin birleşmesi, gelişen kalbin kranial ucundan başlar kaudale doğru uzanır. Kalp tüpleri birleşince, embriyonik kalbin dış tabakası, primordium miyokardiyale, perikardiyal sölom’u saran splanknik mezodermden oluşur. Bu aşamada gelişen kalp ince endotelyal bir tüptür. Endoteliyal tüp, kalbin iç endoteliyal örtüsünü yapar yani endokardiyumu oluşturur. Primordium miyokardiyale, miyokardiyumu oluşturur. Visseral perikardium veya epikardium ise sinus venosus’un dış yüzünden köken alan ve miyokardiyumun üzerine dağılan mezoteliyal hücrelerden farklılaşır. Tübüler kalp uzar, genişleme ve daralmalar oluşur.
Tübüler trunkus arteriozus aorta yaylarının çıktığı sakkus aortikus ile devam eder. Kalp kendi üzerine kıvrılır ve bu şekilde bulboventriküler kıvrımı yapar. Kanalis atriyoventrikülarisin bölmelenmesi; yani primordium atriyale ve ventrikülare oluşumu dördüncü haftanın ortalarında başlar ve beşinci haftanın sonunda tamamlanır. Kalbin primordiumu dört oda içerir:
- Bulbus kordis 2. Ventrikül 3.Atriyum 4.Sinus venozus
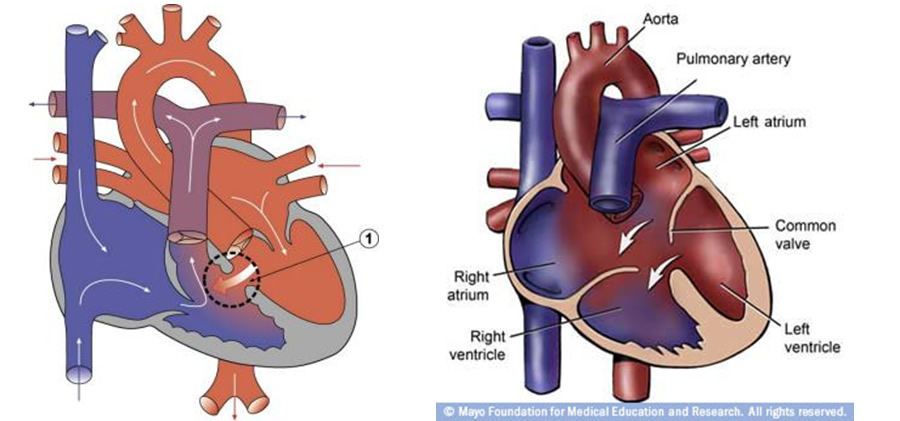
Sinus venozus, primitif atriyumun dorsal duvarının ortasına açılır. Sağ ve sol olmak üzere iki boynuzu vardır. Sol boynuz koroner sinüs olur. Sağ boynuz ise genişler, sağ atriyum duvarıyla birleşerek vena kava superior, vena kava inferioru oluşturur ve plasentadan gelen kanı alır. Dördüncü haftanın sonundan başlayarak primordium atriyale, iki septanın (septum primum ve septum sekundum) oluşumuyla sağ ve sol atriyumlar olarak bölünür. Sol atriyum genişledikçe, primordium vena pulmonalis ve ana dalları sol atrium duvarıyla birleşir ve sonuç olarak 4 pulmoner ven oluşur. Primordium ventrikülare iki septumun (muskuler septum ve membranöz septumun) oluşumuyla sağ ve sol ventriküller olarak bölünür. Kalp dördüncü ve yedinci haftalar arasında dört boşluğa bölünür. Septum aortiko pulmonale ile aorta ve trunkus pulmonalis iki arteriyel kanala ayrılır. Trunkus arteriosus’un bölmelenmesi hemen hemen tamamlandığında, aorta ve trunkus pulmonalis açıklıklarının etrafındaki subendokardiyal dokudaki 3 şişkinlikten valva semilunaris (valva aortika ve valva pulmonalis) gelişmeye başlar. Valva atriyoventrikülaris (valva trikuspidalis ve valva mitralis) benzer olarak AV kanalların çevresindeki dokunun bölgesel çoğalmasından meydana gelir. Kalp büyüdükçe sağa doğru kıvrılır ve erişkin görünümünü alır. Kalbin gelişiminin kritik dönemi fertilizasyondan sonraki 20-50. günlerdir. Bu zamanda normalden herhangi bir sapma, bir veya daha fazla doğumsal kalp defekti oluşturabilir.
Gelişim süresince sağ-sol asimetrisi gösteren vücuttaki ilk yapı kalptir ve bu asimetrinin ilk belirtisi primitif kalp tüpünün öne ve sağa dönüşüdür. Bu da d-loop adını alır. Gelecekteki sol ventrikülü oluşturacak primitif ventrikül de sola doğru değiştirir. Dönüş sonrası kalp, gelişen sağ ventrikülü oluşturmak için solun önüne doğru döner. Kalp sağ yerine sola döndüğünde (l-loop) ventriküllerin normal pozisyonu toraksta yer değiştirir. Morfolojik sağ ventrikül sola, morfolojik sol ventrikül de sağa uzanır. Bu sol-sağ yer değişimi diğer tüm organlarda sol-sağ aks değişimine eşlik eder. Sonuçta situs inversus oluşur. Belirgin olarak birçok situs inversuslu vakada daha sonraki kalp gelişimi normaldir. Buna rağmen normal situslu l-loop’u olan hastalarda ciddi kalp defektleri meydana gelir. Sıklıkla l-loop AV kapak göçünde yetersizliğe neden olur. Bu da çift çıkımlı sol ventrikül ile sonuçlanır. Eğer AV kapak göçü oluşursa sol ventrikül sağ atriyumu ve sol atriyum da sağ ventrikülü takip eder. Bu koşulda eğer konus ve turunkus gelişimi normalse (sağ ventrikül pulmoner artere ve sol ventrikül aortaya açılıyorsa) sonuç izole ventriküler inversiyondur. Daha sık olarak l-loop ventriküloarteriyel bağlantıyı yanlış takip edebilir. Sağ ventrikül aortaya, sol vertrikül de pulmoner artere açılır. Tam sonuç; sol ventrikül, sağ atriyum ve pulmoner arter arasında, sağ ventrikül de sol atriyum ile aorta arasındaki interpozisyonudur. Bu da doğumsal olarak düzeltilmiş büyük arterlerin transpozisyonudur (L-Transpozisyon).
Kardiyak gelişimin en kolay ve en sık kullanılan belirleyicilerinden biri başlangıçtaki kas protein gen ekspresyonu olup mRNA’ya uyan birikim olarak tanımlanır. Bundan başka kontraktil protein geninin düzenlenmesinde çeşitli anahtar faktörler içeren transkripsiyonel faktör tanımlanmıştır. Bunlar kas kreatin kinaz genini düzenleyen MEF-2 (miyosit spesifik arttırıcı faktör 2) olarak bulunmuştur ve birçok kontraktil protein geninin düzenlenmesini içerir. Dört adet mef-2 gen bulunmuştur. Her biri çok sayıda izoformları kodlar. Farede mef-2C döllenmeden 7,5 gün sonra ön splanknik mezodermde ve 11,5 gün sonra kalp tüpünün miyokardiyumunda, venöz sinüste açığa çıkar. mef-2A ve mef-2D 8.günde (kardiyak dönüş safhasında) ve 13,5 güne kadar devam eder. Erken kalp gelişiminde açığa çıkan diğer ailesel faktör GATA faktörüdür. GATA-4’ün açığa çıkması için kontraktil protein genine ihtiyaç vardır ve iskelet kasını oluşturan α miyozin ağır zincirinin gen promotor bölgesini aktive eder. GATA-4 kardiyak spesifitenin belirlenmesinde rol oynar. Bu gen majör iskelet kaslarında normal açığa çıkmayan α miyozin ağır zincir geni olarak kardiyak spesifitenin belirlenmesinde rol oynar. Farede GATA-4 ekspresyonun başlaması 7,5 günde anterior endodermde ve mezodermde olur.
Kalp içinde bu dönemde kaudalde venöz sinüsün sonu ve splanknik mezoderm ile ekspresyon sınırlandırılmıştır. GATA-4, 8. günde kalp tüpü boyunca meydana gelmektedir. Bu ailenin diğer üyeleri kalpte açığa çıkan GATA-5, endokardiyum ve miyokardiyumun her ikisinde, lateral plağın diğer derivelerinde açığa çıkar. Çok uzak ilişkili homeobox genlerinin tersine, gelişen kalbin çevresinde açığa çıkmaktadır. Nkx2.5 ya da Csx olarak adlandırılan iki grup bağımsız homeobox geni gösterilmiştir. Gelişmekte olan kalpte ve erişkin miyokardiyumda açığa çıkar. Daha sonra miyokardiyal tüp ve sinüs venozus boyunca gözlenir. Fonksiyonel rolü kesin olarak bilinememektedir. Diğer homeobox genleri kalp içinde açığa çıkar. Bunlar Msx-1, Msx-2, MHox genleridir. Msx-2 kuşlarda, gelişen miyokardiyum alanında gözlenir ve iletim sisteminin gelişimine katkıda bulunur. Msx-1 endokardiyal yastık dokularında açığa çıkar ve bu dokudaki mezenşimal hücre populasyonunun oluşmasında rol alır. Atriyoventriküler bağlantıda kenar doku mezenkimi, endokardiyum hücrelerinin tabaka halinde diziliminden oluşur. Msx-1’in ekspresyonu mezenşimin gelişiminin devamında gözlenir. MHox kas kreatin kinaz geninin düzenlenmesinde rol oynayan adenin-timinden zengin sekans motifine bağlanmada bir faktör olarak tanımlanmıştır. Fetal hayattan erişkin döneme kadar kardiyak kas geninin düzenlenmesinde rol oynar.
Dominant fonksiyonel özellik olarak ayrılan miyokardiyum, erken kalp tüpünden daha ileri dönemde yavaş impuls üretir. Pacemaker aktivitesi erken dönemde kardiyak tüp içinde vardır. İmpulsların yavaş üretilmesi tübüler kalpte karakteristik olan peristaltoid form kontraksiyonlara yol açar. Kalbin üst kısımları hızlı, alt kısımları yavaş kontraksiyonlarla karakterizedir. Kısa zamanda hızlı miyozin formları (α–MHC) ve sarkoplazmik retikulum kalsiyum ATP az (SERCA-2), uzun zamanda da yavaş miyozin formları (β–MHC) ve az miktarda SERCA-2 açığa çıkar. Embriyonik ventrikül, trabeküler ve kompakt miyokardiyal komponentler olarak ayrılmışlardır. Erken ventrikül miyokardiyumunun hepsi trabekülerdir. Majör kardiyak “gap junctional protein connexin 43” ventriküler trabekülasyonlarda en yüksek seviyede açığa çıkmaktadır. SERCA-2 trabeküler komponentte düşük olmaktadır. Benzer mRNA seviyeleri karşılaştırıldığında kompakt miyokardiyumda oldukça düşüktür. Connexin 40 iletim sisteminin bir kısmında baskın gap junctional proteindir. Kontraktil komponentler tek genle kodlanmaz. Bu multigen ailenin her bir üyesinin farklı protein izoformları vardır. Bunlardan biri olan miyozinin, iki adet miyozin ağır zinciri iki adet miyozin hafif zincir tipleri vardır. (Miyozin hafif zincir 1 (MLC1), miyozin hafif zincir 2 (MLC2)). Bu proteinlerin her birinin çok sayıda izoformu vardır ve tek genle kodlanarak ayrılırlar. Kardiyak kasta açığa çıkan iki miyozin ağır zincir izoformları (α-MHC ve β-MHC) birbirine yakın bağlı genle kodlanırlar. α-MHC en yüksek oranda kalbin giriş yolunda, β-MHC ise kalbin çıkış yolunda bulunur.
Atriyumlarda α izoform ventriküllerde β izoform bulunmaktadır. β-MHC atriyoventriküler bağlantı sınırında açığa çıkmaktadır. Fakat yeni oluşan kalpte atriyoventriküler sınırda olmamaktadır. Küçük memelilerde ilk haftada ventriküllerde açığa çıkan β-MHC gelişim esnasında α-MHC ile yer değiştirir. Büyük memelilerde ve insanlarda β-MHC ventriküler kaslarda baskın izoform olarak açığa çıkıp gelişimde ve ergenlikte devam etmektedir. Ayrılan genlerin kodladığı yetişkinlerdeki miyozin hafif zincirin atriyal izoformları MLC1A, MLC2A ve yetişkin ventriküler izoformları MLC1V, MLC2V’dir.Tip 1 miyozin hafif zincirleri başlangıçta kardiyak tüp boyunca açığa çıkar. Atriyal tip 2 miyosin hafif zincir konsantrasyonu kalbin giriş yollarından çıkış yollarına doğru azalır. Ventriküler Tip 2 miyozin hafif zinciri buna rağmen bölgesel sınırlanmış ekspresyon paterni gösterir. MLC2A ve MLC2V miyokardiyumda, kalbin giriş yollarında, atriyoventriküler kanalda, kalbin çıkış yollarında erişkin hayata kadar eksprese olurlar. İnce filament protein olan α aktinin üç izoformu kalp kasında açığa çıkmaktadır ve ayrı genlerle kodlanmaktadır. Đnsan kalbinde α iskelet aktini ilk papiller kaslarda bulunmuştur. Diğer izoformlarının gelişimsel değişimleri henüz aydınlatılamamıştır. İnce filament proteinleri olan troponin kompleksin troponin Ι, troponin T, ve troponin C olmak üzere üç izoformu vardır. Đki farklı troponin C geni vardır. Hızlı iskelet ve yavaş iskelet / kardiyak izoformları kodlar. Yavaş kardiyak izoform her iki atriyal ve ventrikül miyokardında gelişimde ve erişkinlikte açığa çıkar. Troponin Ι’nın iki geni gelişen kalpte açığa çıkar. Bunlar erişkin kalp izoformu (TnΙc) ve yavaş iskelet kas izoformunu kodlar. Erişkin atriyal ve ventriküler myokardiyumda bulunan tek izoform kardiyak izoformdur. Tek genle açığa çıkan troponin T, kalbin gelişimi boyunca eksprese olmaktadır.